| IV CONGRESO VIRTUAL HISPANO AMERICANO DE ANATOMÍA PATOLÓGICA |
|
CONTENIDO |
|
|
|
|
|
IMÁGENES | ||||||||
|
Fig. 7
Fig. 8
Fig. 9
Fig. 10
Fig. 11
Fig. 12 |
|
|
|
INTRODUCCIÓN | |
| La apolipoproteína D (apo D) es una glicoproteína, con numerosas isoformas, que forma parte de las partículas de lipoproteína presentes en el plasma (1). Pertenece a la extensa familia de las lipocalinas, transportadoras de pequeños ligandos hidrofóbicos (2, 3). La apo D se sintetiza en una gran variedad de tejidos como el hígado, intestino, páncreas, riñón, placenta, glándulas adrenales, bazo y cerebro (2, 4). Se expresa en gran variedad de tejidos (5-7). No se puede considerar que el ARNm de la apo D sea tejido-específico, hasta el momento los lugares de mayor expresión encontrados han sido, junto con el hígado, el sistema nervioso central (SNC) y el sistema nervioso periférico (SNP). Fundamentalmente encontramos su ARNm en fibroblastos, a menudo asociados con vasos sanguíneos (2, 4, 6, 7). Varios trabajos parecen señalar que distintos tipos de células gliales y posiblemente neuronas, pudieran proveer al SNC de apo D (8, 9). Se ha visto como las hormonas esteroides son capaces de regular la secreción de apo D “in vitro” (10, 8). También se han identificado distintos elementos reguladores en la región promotora del gen de la apo D (11). Debido a la multitud de ligandos y a la gran variedad de lugares de síntesis, se cree que la apo D podría ser una proteína multiligando y multifuncional, importante en la fisiología celular y de función variable según el lugar de síntesis. A nivel sistémico y en asociación con la lecitin colesterol acetiltransferasa (LCAT), la apo D forma un complejo que actúa de forma activa retirando colesterol de los tejidos periféricos y transportándolo por el plasma hasta el hígado para su catabolismo (12). Existen varias líneas de trabajo que sugieren un papel para la apo D en las situaciones de degeneración y regeneración del SN, por ejemplo, en la enfermedad de Niemman – Pick tipo C (NPC) (13, 14) y en la enfermedad de Alzheimer (EA) (15, 16). Existen también evidencias de la importancia del papel de la apo D en la redistribución lipídica después de la injuria experimental de un nervio (5, 17), lo que sugiere que esta proteína puede representar un papel muy importante en el transporte lipídico a nivel periférico. También se ha visto como la presencia de apo D aumenta durante el envejecimiento en el cerebelo humano, presentando las neuronas inmunopositivas unas características morfológicas que hacen pensar en la apo D como en un posible marcador de necrobiosis
(18). La apoD puede estar relacionada con el crecimiento mediado por esteroides en algunas líneas de tumores
(19). La vitamina D3 y otras hormonas esteroideas parecen ser inductoras de la expresión de apoD en algunos carcinomas de mama
(20,21).
|
||
|
|
MATERIAL Y MÉTODOS | |
Para la realización del presente estudio se utilizaron 43 autopsias procedentes del Servicio de Anatomía Patológica del Hospital Central de Asturias. En las muestras, incluidas en parafina, se realizaron cortes de 10 mm de grosor sobre los que se efectuó una técnica inmunohistoquímica para la apo D (anticuerpo cedido por el Dr. Carlos López-Otín del Área de Bioquímica de la Universidad de Oviedo) que fue visualizada en un microscopio de campo claro mediante el cromógeno diaminobencidina (DAB). Se estudiaron los vasos subpiales, corticales, subcorticales, periventriculares y lenticulares mediante estimación subjetiva. Se valoraron los siguientes cambios: engrosamiento parietal por capas, pérdida celular por capas, fibrosis por capas, presencia o no de lagunas perivasculares, presencia o no de macrófagos, gliosis y material proteináceo perivascular. Se hizo un estudio descriptivo y analítico utilizando el paquete estadístico SPSS para Windows. Se utilizaron los análisis estadísticos básicos tabulando los datos y transformando y reagrupando las variables en los casos necesarios. Inicialmente se hizo una descripción de frecuencias, para pasar luego a un análisis exploratorio de datos. Posteriormente se cruzaron las variables en tablas simples de contingencia. En los cruces de variables por medio de estas tablas, pudimos ver las relaciones entre variables y el sentido de tal relación, comprobando el aspecto inferencial con la significación del c2, considerando estadísticamente significativas las cifras de p > 0,05. |
||
|
|
RESULTADOS | |
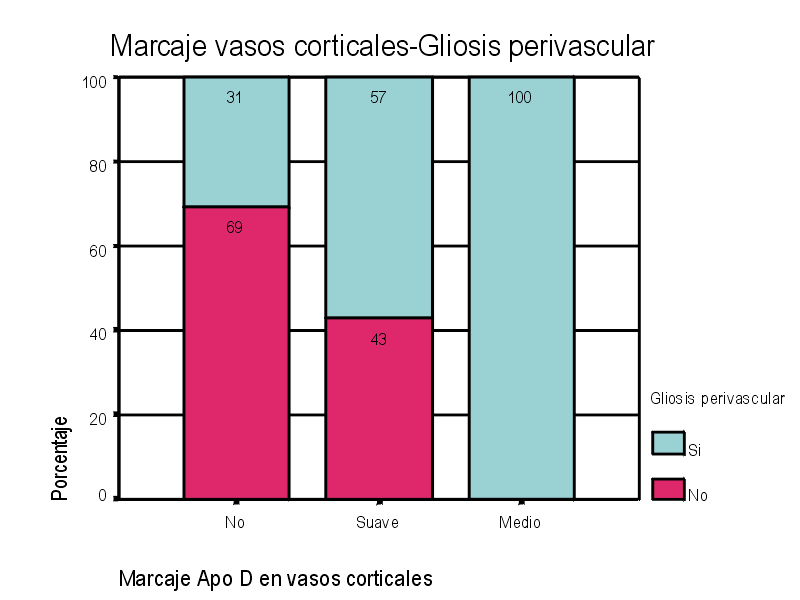
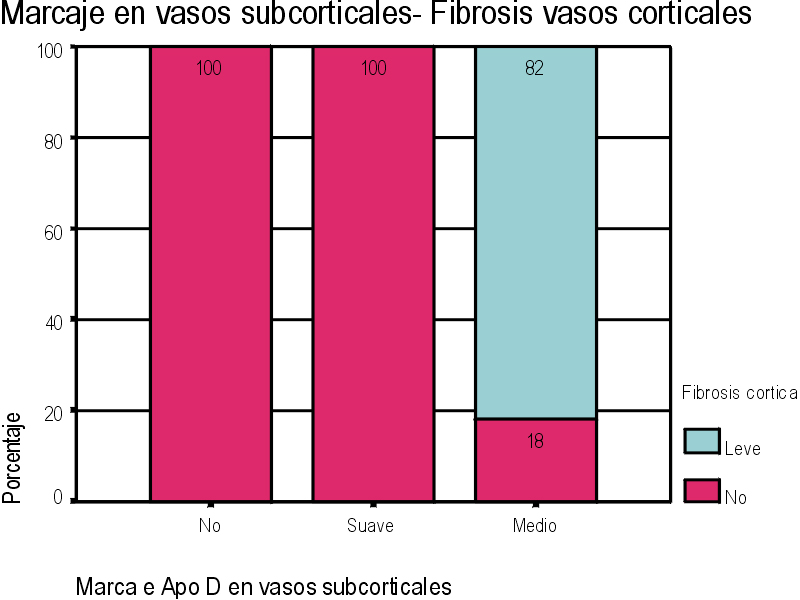
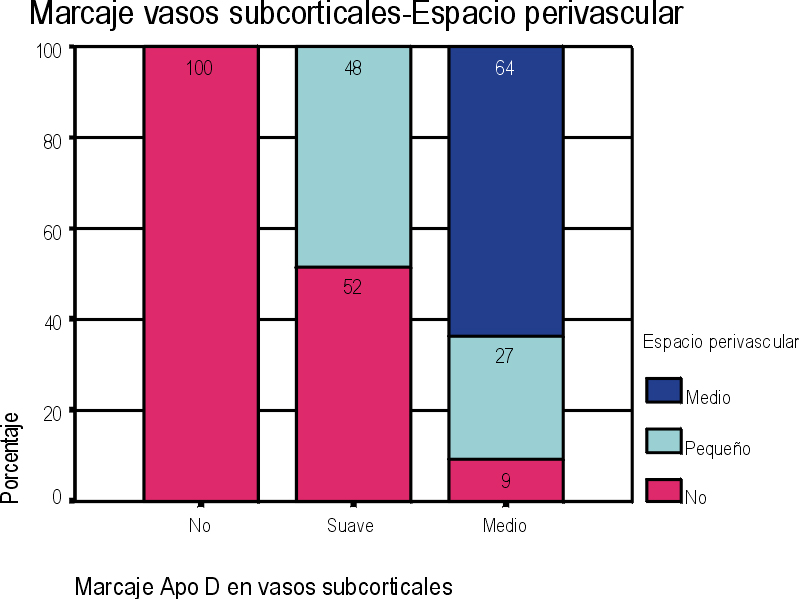
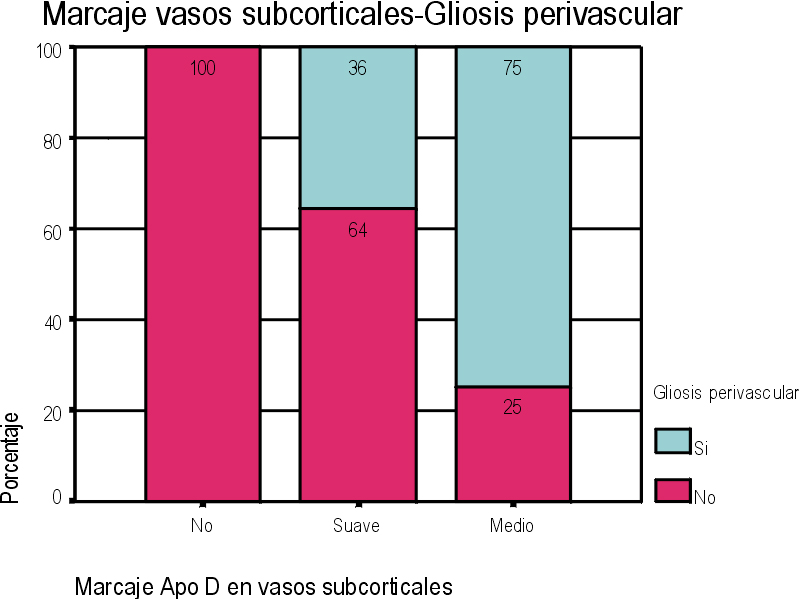
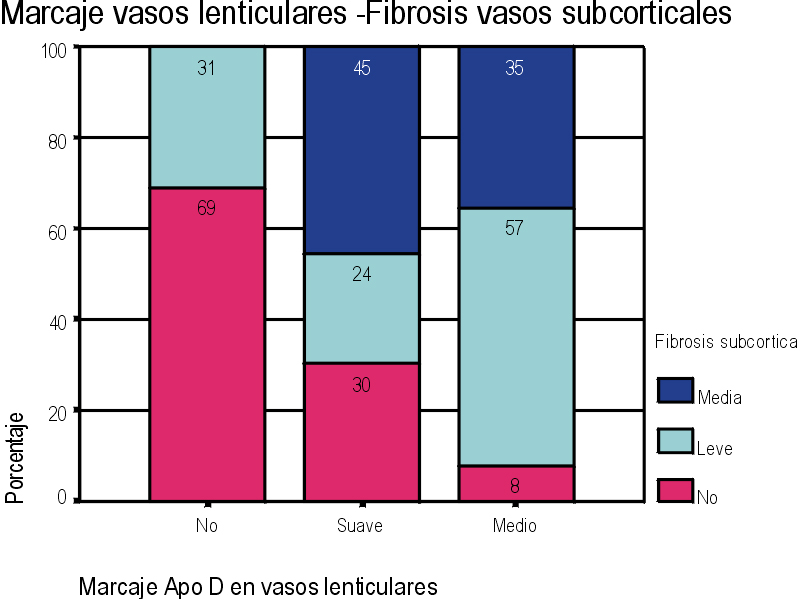
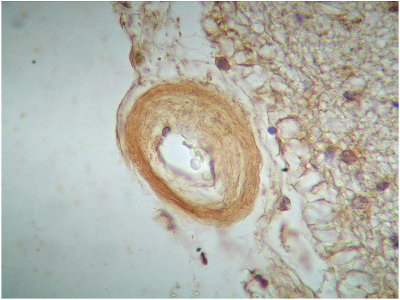
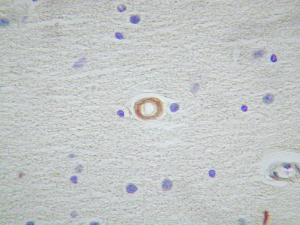
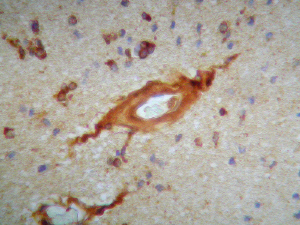
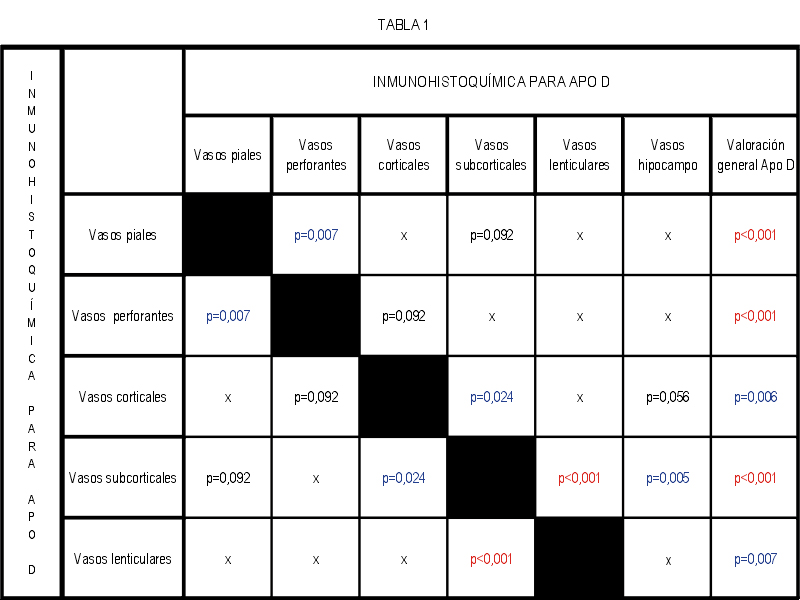
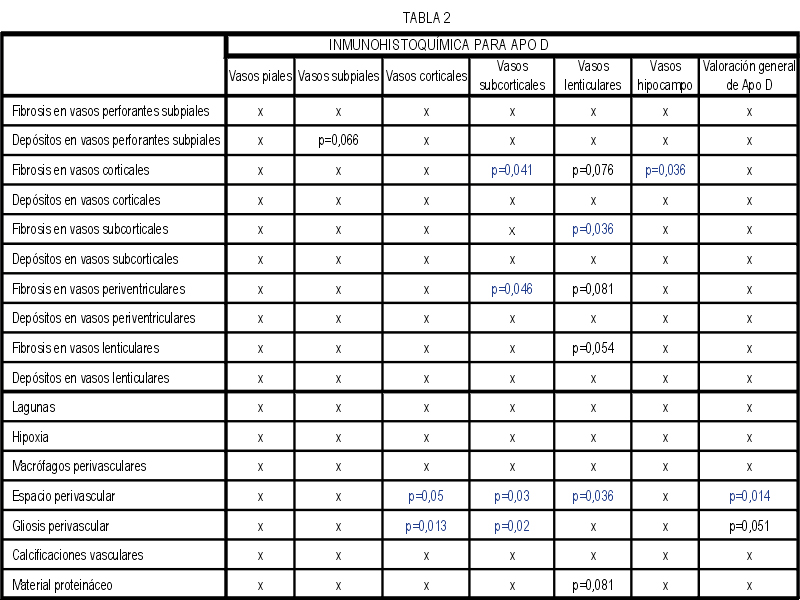
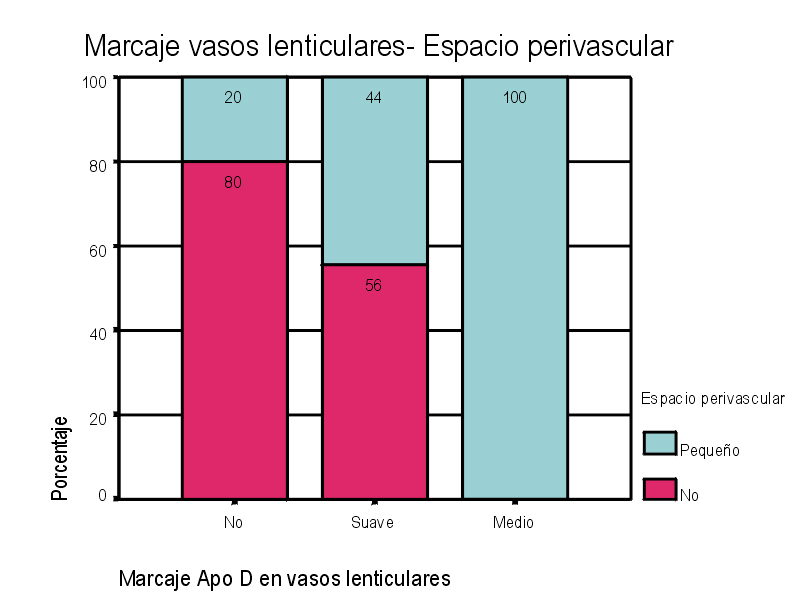
El marcaje IH para apoD se observa al microscopio óptico con una coloración que va del color marrón claro al color marrón oscuro en función del grado de depósito de apoD. No existe relación directa entre el marcaje IH(+) por vaso y el marcaje IH (+) en número de vasos afectados. Existe cierto grado de correlación entre la inmunotinción positiva para vasos piales y subpiales, en ambos tipos de vasos el marcaje se presenta de forma homogénea (Figura 1 y Figura 2). Los vasos corticales se muestran inmunopositivos en una proporción inferior a los vasos piales y subpiales (Figura 3). Los depósitos son en placas, aunque se observaron casos de afectación generalizada. Los depósitos son preferentemente granulares, aunque también se observaron depósitos mixtos. La afectación por capas de la pared tiene tendencia a ser global aún en los casos de depósitos en placas. En la línea de unión cortical-subcortical existe un aumento de la inmunotinción. En el caso de los vasos subcorticales, se pueden observar zonas de inmunotinción positiva intercaladas con zonas inmunonegativas: al inicio los depósitos son lineales, fibrilares, con afectación variable de las capas; al aumentar la tinción, los depósitos adquieren una apariencia más granular, mantienen la distribución en placas pero aumentan el número de capas vasculares afectadas (Figura 4). Los vasos lenticulares presentan al inicio depósitos no homogéneos, con predominio en adventicia y apariencia fibrilar. Al aumentar la tinción los depósitos se hacen más generalizados y ocupan toda la pared, adquiriendo un aspecto mixto, granular y fibrilar. El comportamiento de los vasos del hipocampo fue similar al observado en el resto de la corteza cerebral. Se relacionaron entre si dos a dos los valores obtenidos para el marcaje IH de apo D en los distintos vasos cerebrales estudiados (Figura 5). También se enfrentaron los hallazgos histológicos compatibles con la arteriosclerosis con los valores obtenidos para la presencia de apo D en los vasos cerebrales (Figura 6). Se estableció en ambos casos la existencia o no de significación estadística. Los hallazgos histológicos más significativos, en relación con el inmunomarcaje positivo de los vasos para apo D, resultaron ser la fibrosis, la gliosis perivascular y el espacio perivascular (Gráficas 1-8). |
||
|
|
DISCUSIÓN | |
Hay una evidencia creciente que indica que la temprana alteración del metabolismo lipídico durante la injuria puede jugar un papel crítico en la muerte neuronal y en la actividad de las células gliales (22-27). Se postula que debido a la excitotoxicidad se produce un aumento de fosfolipasa A2, lo que produce descomposición de fosfolípidos que se vuelven perjudiciales para la neurona. Excesivo acúmulo de ácidos grasos libres, lisofosfolípidos y ácido lisofosfatídico se relacionó con la alteración de la respiración mitocondrial y la homeostasis neuronal, (27, 28, 24, 29) induciendo necrosis y apoptosis en neuronas del hipocampo (30). El ácido araquidónico, abundante en los fosfolípidos del cerebro, actúa como un mensajero retrógrado de la transmisión glutamatérgica (31), como un substrato para la ciclooxigenasa y la lipooxigenasa y también probablemente como un regulador de la expresión genética (24) pudiendo producir directa o indirectamente efectos en el estado del cerebro (32-35). Eventualmente, debido a esa intensa desaparición fosfolipídica puede también existir lisis osmótica de la membrana celular, produciéndose entonces directamente grandes acúmulos de lípidos de membrana libres que pueden ser oxidados en aldehídos tóxicos que induzcan numerosas disfunciones cerebrales
(33, 26). El retraso en la recaptación de lipopartículas puede también ser altamente destructor de la supervivencia neuronal ya que ha sido demostrado que las LDL y HDL oxidadas promueven la muerte neuronal
(36). Por lo tanto, cuando patológica o experimentalmente existe lesión neuronal la inmediata protección de la neurona, mediante el empaquetado y la recaptura de lípidos, puede ser esencial para disminuir el efecto letal de la excitotoxicidad. Y ese papel parece que está en posible relación con la apolipoproteína D por nosotros estudiada.
|
||
|
|
NOTAS AL PIE DE PÁGINA | |
Correspondencia: E. del Valle E. Universidad de Oviedo. Oviedo, España. mailto:jtolivia@correo.uniovi.es Patrocinadores: Este trabajo ha sido financiado por el proyecto del
F.I.S. 99/1316
|
||